Inicio  ÓRGANO INTERNO DE CONTROL Ciencia hoy El ritmo de la evolución de las cícadas del género Ceratozamia
ÓRGANO INTERNO DE CONTROL Ciencia hoy El ritmo de la evolución de las cícadas del género Ceratozamia
El ritmo de la evolución de las cícadas del género Ceratozamia
Jorge González-Astorga, Anwar Medina-Villarreal y Alejandro Espinosa de los Monteros
 En 1986 el cicadólogo Dennis Stevenson propuso algunas ideas o hipótesis sobre la evolución del género Ceratozamia. Sin embargo, no fue sino hasta ahora, pasados treinta y tres años, que se rescataron y se pusieron a prueba. Para ello, un punto de partida fundamental fue el análisis de caracteres fenotípicos sobre un marco filogenético-molecular, temporalmente explícito. Esto permitió inferir que la evolución ocurrió a ritmos diferenciales dentro de los dos principales linajes y sugiere que los mecanismos de especiación fueran moldeados por procesos de selección-adaptación, posiblemente asociados al estrés hídrico; y potenciados por procesos estocásticos como deriva génica-endogamia asociados a los tamaños poblacionales pequeños característicos de este grupo de plantas.
En 1986 el cicadólogo Dennis Stevenson propuso algunas ideas o hipótesis sobre la evolución del género Ceratozamia. Sin embargo, no fue sino hasta ahora, pasados treinta y tres años, que se rescataron y se pusieron a prueba. Para ello, un punto de partida fundamental fue el análisis de caracteres fenotípicos sobre un marco filogenético-molecular, temporalmente explícito. Esto permitió inferir que la evolución ocurrió a ritmos diferenciales dentro de los dos principales linajes y sugiere que los mecanismos de especiación fueran moldeados por procesos de selección-adaptación, posiblemente asociados al estrés hídrico; y potenciados por procesos estocásticos como deriva génica-endogamia asociados a los tamaños poblacionales pequeños característicos de este grupo de plantas.
Palabras clave: Fósiles vivientes, México, pedomorfosis, variación foliar, Zamiaceae.
La evolución o descendencia con modificación, es un proceso de cambios de la composición genética y fenotípica de las especies espacio-temporalmente. Este cambio puede ser continuo, lento y gradual, o bien discontinuo, rápido y episódico (Gould 2002). Para que la evolución ocurra, se requiere suficiente variación genética que permita la acción diferencial de la selección natural, y en consecuencia la adaptación de los organismos (Fisher 1958). Simultáneamente, en mayor o menor grado, actúan fuerzas evolutivas que propician la mezcla entre poblaciones ais-ladas (flujo génico); o bien otras que contribuyen a su aislamiento (deriva génica y endogamia; Wright 1931). En el cambio evolutivo rápido es necesario que los mecanismos que originan variabilidad genética de novo, como la mutación y la re-combinación genética, tengan efectos di-ferenciales significativos sobre el fenotipo. Al respecto, Waddington (1968) propuso que el epigenotipo, es decir la interacción entre el genotipo y el fenotipo, está relacionado con procesos que ocurren durante la biología del desarrollo u ontogenia de los organismos. La duplicación de juegos completos de cromosomas o poliploidía es un ejemplo de evolución rápida. En plantas se estima que aproximadamente el 50% de las especies se han originado a través de este mecanismo (Grant 1989). Existen evidencias suficientes de que en la evolución temprana de las angiospermas estuvo relacionada con eventos de duplicación del genoma completo, y se ha demostrado que la formación de poliploides tiene factores causales epigenéticos (De Bodt et al. 2005). Otro proceso genético que potencia la rápida especiación es la hibridación, por la fusión exitosa de genomas de especies filogenéticamente cercanas (Riese-berg 1997).
 Las especies en la naturaleza se pueden agrupar por su historia común (filogenia), por el ambiente o área geográfica que habitan (biogeografía) y por sus similitudes en atributos morfológicos vegetativos y reproductivos (fenética). En este contexto, un grupo de especies vegetales muy in-teresantes son las cícadas, ya que estas “son para el reino vegetal, lo que los dinosaurios son para el animal, porque representan la culminación de las dinastías dominantes de la era Mesozoica” (sensu Ward 1900). Aunque su origen data de hace 250 millones de años, las especies contemporáneas surgieron alrededor de hace 20 millones de años (Nagalingum et al. 2011) o aún menos (Calonje et al. 2019a).
Las especies en la naturaleza se pueden agrupar por su historia común (filogenia), por el ambiente o área geográfica que habitan (biogeografía) y por sus similitudes en atributos morfológicos vegetativos y reproductivos (fenética). En este contexto, un grupo de especies vegetales muy in-teresantes son las cícadas, ya que estas “son para el reino vegetal, lo que los dinosaurios son para el animal, porque representan la culminación de las dinastías dominantes de la era Mesozoica” (sensu Ward 1900). Aunque su origen data de hace 250 millones de años, las especies contemporáneas surgieron alrededor de hace 20 millones de años (Nagalingum et al. 2011) o aún menos (Calonje et al. 2019a).
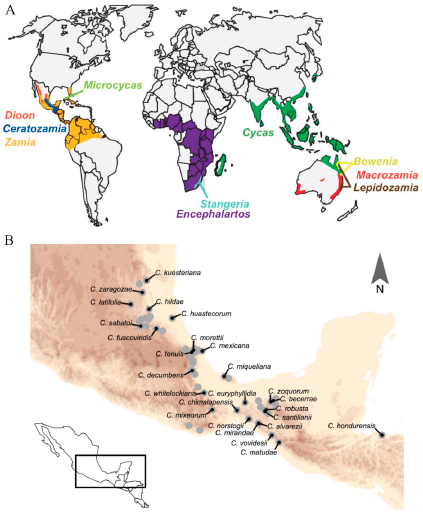
Actualmente, las 355 especies existentes de cícadas están repartidas en diez géneros distribuidos en las zonas tropicales de los cuatro  continentes: Cycas L. en Asia; Bowenia Hook. ex Hook. f., Macrozamia Miq., Lepidozamia Regel y Cycas L. en Oceanía; Encephalartos Lehm. y Stangeria T. Moore en África; y por último Microcycas (Miq.) A. DC., Dioon Lindl., Zamia L., y Ceratozamia Brongn. en América (Figura 1A). En cuanto a la riqueza de especies, México ocupa el segundo lugar con 61 spp. solo superado por Australia con 78 spp. (Calonje et al. 2019b). En México se distribuyen tres géneros: Dioon con 17 spp, Zamia con 15 spp y Ceratozamia con 29 spp. De estas especies, la mayoría son endémicas (i.e., están restringidas) en regiones bien definidas, y con distribuciones geográficas que no se solapan. De los tres géneros mexicanos, el género Ceratozamia es el más rico en especies y con la mayor diversidad morfológica. Sus especies se distribuyen en tipos de vegetación selváticos y bosques húmedos, desde el norte del Eje Neovolcánico Transversal a lo largo de la Sierra Madre Oriental, hasta Honduras (Figura 1B). En las plantas adultas de este género se ha registrado una amplia diversidad foliolar, desde formas estrechas como en C. norstogii D.W. Stev., hasta formas anchas como en C. euryphyllidia Vázq.-Torres, Sabato y D.W. Stev. (Figura 2).
continentes: Cycas L. en Asia; Bowenia Hook. ex Hook. f., Macrozamia Miq., Lepidozamia Regel y Cycas L. en Oceanía; Encephalartos Lehm. y Stangeria T. Moore en África; y por último Microcycas (Miq.) A. DC., Dioon Lindl., Zamia L., y Ceratozamia Brongn. en América (Figura 1A). En cuanto a la riqueza de especies, México ocupa el segundo lugar con 61 spp. solo superado por Australia con 78 spp. (Calonje et al. 2019b). En México se distribuyen tres géneros: Dioon con 17 spp, Zamia con 15 spp y Ceratozamia con 29 spp. De estas especies, la mayoría son endémicas (i.e., están restringidas) en regiones bien definidas, y con distribuciones geográficas que no se solapan. De los tres géneros mexicanos, el género Ceratozamia es el más rico en especies y con la mayor diversidad morfológica. Sus especies se distribuyen en tipos de vegetación selváticos y bosques húmedos, desde el norte del Eje Neovolcánico Transversal a lo largo de la Sierra Madre Oriental, hasta Honduras (Figura 1B). En las plantas adultas de este género se ha registrado una amplia diversidad foliolar, desde formas estrechas como en C. norstogii D.W. Stev., hasta formas anchas como en C. euryphyllidia Vázq.-Torres, Sabato y D.W. Stev. (Figura 2). 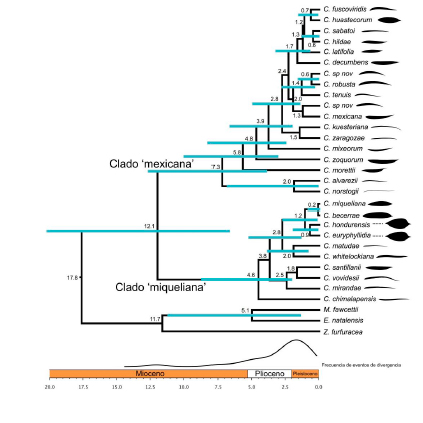 Por el contrario, la variación es mínima en las plántulas de todas las especies (Figura 3). Los foliolos de las plántulas son anchos, y tan similares que hasta a un experto le resultaría difícil distinguir a que especie pertenece. Al respecto y con la idea de dar respuesta al origen de este género, Stevenson et al. (1986) propusieron que la divergencia fenotípica en Ceratozamia habría ocurrido en dos trayectorias. Una ruta consiste en especies que perpetúan las formas juveniles hasta la madurez, mientras que la otra ruta consiste en especies con cambios ontogenéticos que las diferencian del estado juvenil del maduro. Para la segunda ruta sugirieron que fenotipos similares en estado adulto, habrían evolucionado de forma independiente bajo presiones ambientales similares. Con base en lo anterior, Medina-Villarreal et al. (2019) pusieron a prueba las ideas propuestas por Stevenson et al. (1986) y determinaron que la evolución de las especies de Ceratozamia es explicada por un efecto combinado por la interacción entre historia filogenética y el ambiente. Sus resultados sugieren que existe una importante correlación fenotipo-ambiente, relacionado a ritmos dife-renciales en cada evento de cladogénesis (Figura 4). A este modo de evolución con diferentes ritmos o tasas se le conoce como evolución puntuada, donde los cam-bios ocurren de forma rápida y posterior al evento de especiación o de aislamiento reproductivo. En este proceso, sucede que una rama hermana mantiene el carácter ancestral y entra en un periodo de estasis morfológica, mientras que la otra rama cambia (Oakley et al. 2005). El cambio en una de las ramas a menudo está asociado a cambios de nicho (Gutiérrez-Ortega et al. 2018), los cuales ocurren comúnmente en las radiaciones adaptativas influenciadas por el ambiente (Good-Avila et al. 2006). Para evidencia de lo anterior ponen de ejemplo pares de especies hermanas: unas paedomorficas y otras peramórficas (Figura 5). Las primeras, corresponden a aquellas que mantienen la forma juvenil hasta la adultez, mientras en las segundas corresponden a aquellas que son más distintas que lo esperado
Por el contrario, la variación es mínima en las plántulas de todas las especies (Figura 3). Los foliolos de las plántulas son anchos, y tan similares que hasta a un experto le resultaría difícil distinguir a que especie pertenece. Al respecto y con la idea de dar respuesta al origen de este género, Stevenson et al. (1986) propusieron que la divergencia fenotípica en Ceratozamia habría ocurrido en dos trayectorias. Una ruta consiste en especies que perpetúan las formas juveniles hasta la madurez, mientras que la otra ruta consiste en especies con cambios ontogenéticos que las diferencian del estado juvenil del maduro. Para la segunda ruta sugirieron que fenotipos similares en estado adulto, habrían evolucionado de forma independiente bajo presiones ambientales similares. Con base en lo anterior, Medina-Villarreal et al. (2019) pusieron a prueba las ideas propuestas por Stevenson et al. (1986) y determinaron que la evolución de las especies de Ceratozamia es explicada por un efecto combinado por la interacción entre historia filogenética y el ambiente. Sus resultados sugieren que existe una importante correlación fenotipo-ambiente, relacionado a ritmos dife-renciales en cada evento de cladogénesis (Figura 4). A este modo de evolución con diferentes ritmos o tasas se le conoce como evolución puntuada, donde los cam-bios ocurren de forma rápida y posterior al evento de especiación o de aislamiento reproductivo. En este proceso, sucede que una rama hermana mantiene el carácter ancestral y entra en un periodo de estasis morfológica, mientras que la otra rama cambia (Oakley et al. 2005). El cambio en una de las ramas a menudo está asociado a cambios de nicho (Gutiérrez-Ortega et al. 2018), los cuales ocurren comúnmente en las radiaciones adaptativas influenciadas por el ambiente (Good-Avila et al. 2006). Para evidencia de lo anterior ponen de ejemplo pares de especies hermanas: unas paedomorficas y otras peramórficas (Figura 5). Las primeras, corresponden a aquellas que mantienen la forma juvenil hasta la adultez, mientras en las segundas corresponden a aquellas que son más distintas que lo esperado 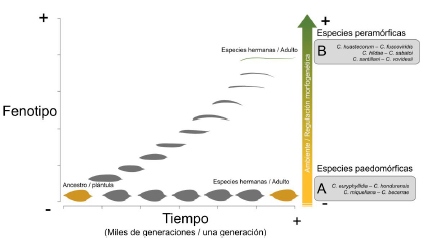 por su distancia filogenética. Se infiere que el motor de cambio fenotípico pudo haber estado relacionado con la intensidad de los cambios climáticos remotos. De forma concluyente, Medina-Villarreal et al. (2019) presentan lo que ellos nombran “escenario onto-filogenético” del género Ceratozamia, el cual es descrito con dos posibles resultados. Por un lado, en la rama de cambio rápido, se infiere que el ambiente actúo como régimen (factor) selectivo en las poblaciones ancestrales (con base en la actual correlación fenotipo-ambiente: la causa próxima) y dirigió la transformación ontogenética durante cientos de generaciones. Esta situación condujo al cambio fenotípico en los linajes (causa última), por lo que este podría ser recuperado en el desarrollo de una generación (i.e. ontogenia) y la historia evolutiva de las especies (i.e. filogenia). En cambio, en las especies donde el ambiente se mantuvo constante, éstas retuvieron los caracteres de los ancestros sin cambio sustantivo en el tiempo (Figura 5). Este mecanismo evolutivo explica en parte el hecho de por qué especies filogenéticamente distantes sean más similares que otras más cercanas. Esto sugiere que, bajo un patrón de evolución convergente de caracteres fenotípicos, como es el caso en el género Ceratozamia (y recientemente encontrado también en Zamia; Calonje et al. 2019a) se enmascaren las relaciones naturales de las especies cuando se usan métodos no-filogenéticos para su reconocimiento (o rechazo), lo cual es importante en la sistemática de este carismático grupo de plantas. En consecuencia, esto nos conduce a promover estudios exhaustivos a nivel especifico y poblacional con filogenias robustas (multi-loci, con varias regiones de ADN) que incluyan análisis de reconstrucción de caracteres, y que permitan entender los procesos que dieron origen a los patrones de diversidad actual. Referencias Calonje M., Meerow A.W., Griffith.
por su distancia filogenética. Se infiere que el motor de cambio fenotípico pudo haber estado relacionado con la intensidad de los cambios climáticos remotos. De forma concluyente, Medina-Villarreal et al. (2019) presentan lo que ellos nombran “escenario onto-filogenético” del género Ceratozamia, el cual es descrito con dos posibles resultados. Por un lado, en la rama de cambio rápido, se infiere que el ambiente actúo como régimen (factor) selectivo en las poblaciones ancestrales (con base en la actual correlación fenotipo-ambiente: la causa próxima) y dirigió la transformación ontogenética durante cientos de generaciones. Esta situación condujo al cambio fenotípico en los linajes (causa última), por lo que este podría ser recuperado en el desarrollo de una generación (i.e. ontogenia) y la historia evolutiva de las especies (i.e. filogenia). En cambio, en las especies donde el ambiente se mantuvo constante, éstas retuvieron los caracteres de los ancestros sin cambio sustantivo en el tiempo (Figura 5). Este mecanismo evolutivo explica en parte el hecho de por qué especies filogenéticamente distantes sean más similares que otras más cercanas. Esto sugiere que, bajo un patrón de evolución convergente de caracteres fenotípicos, como es el caso en el género Ceratozamia (y recientemente encontrado también en Zamia; Calonje et al. 2019a) se enmascaren las relaciones naturales de las especies cuando se usan métodos no-filogenéticos para su reconocimiento (o rechazo), lo cual es importante en la sistemática de este carismático grupo de plantas. En consecuencia, esto nos conduce a promover estudios exhaustivos a nivel especifico y poblacional con filogenias robustas (multi-loci, con varias regiones de ADN) que incluyan análisis de reconstrucción de caracteres, y que permitan entender los procesos que dieron origen a los patrones de diversidad actual. Referencias Calonje M., Meerow A.W., Griffith.

Figuras
Figura 1. Distribución actual de los géneros de Cycadales. B. Distribución de las especies del género Ceratozamia. Los puntos grises representan el total de poblaciones, y los puntos negros representan la población muestreada para el análisis filogenético. (A. Imagen tomada y modificada de Nagalingum et al. 2011. B. Imagen tomada y modificada de Medina-Villarreal et al. 2019).
Figura 2. Diversidad foliolar de algunas especies Ceratozamia (Fotografías: Anwar Medina-Villarreal).
Figura 3. Ejemplos de plantas del género Ceratozamia. A. Adulto de C. mexicana. B. Plántula de C. mexicana. C. Megaestróbilo de C. tenuis. D. Microstróbilo de C. tenuis. E. De izquierda a dere-cha, hojas de C. tenuis, C. mexicana, C. decumbens, C. morettii. (Fotografías: Anwar Medina-Villarreal).
Figura 4. Filogenia molecular calibrada temporalmente con fósiles (cronograma) de las especies del género Ceratozamia. Genes utilizados: nrITS, matK, rbcL, rpoC1, psbK/I y atpF/H. La edad estimada de cladogénesis se indica en cada nodo. (Imagen tomada y modificada de Medina-Villarreal et al. 2019).
Figura 5. Diagrama onto-filogenético de la evolución del genero Ceratozamia. En el eje Y el fenotipo va desde mayor a menos similitud morfológica. Paralelo al eje Y, un vector de intensidad ambiental/regulación morfogenética) como fuerza de transformación fenotípica. En el eje X se representa el tiempo en escala de generaciones. (Imagen tomada y modificada de Medina-Villarreal et al. 2019).
